The measles virus (MV), with scientific name Morbillivirus hominis, is a single-stranded, negative-sense, enveloped, non-segmented RNA virus of the genus Morbillivirus within the family Paramyxoviridae. It is the cause of measles. Humans are the natural hosts of the virus; no animal reservoirs are known to exist.
| Morbillivirus hominis | |
|---|---|

| |
| Measles morbillivirus electron micrograph | |
| Virus classification | |
| (unranked): | Virus |
| Realm: | Riboviria |
| Kingdom: | Orthornavirae |
| Phylum: | Negarnaviricota |
| Class: | Monjiviricetes |
| Order: | Mononegavirales |
| Family: | Paramyxoviridae |
| Genus: | Morbillivirus |
| Species: | Morbillivirus hominis
|
| Synonyms[1] | |
|
Measles virus (MV) | |
Disease
editThe virus causes measles, a highly contagious disease transmitted by respiratory aerosols that triggers a temporary but severe immunosuppression. Symptoms include fever, cough, runny nose, inflamed eyes and a generalized, maculopapular, erythematous rash and a pathognomonic Koplik spot seen on buccal mucosa opposite to lower 1st and 2nd molars. The virus is spread by coughing and sneezing via close personal contact or direct contact with secretions.[2]
Replication cycle
edit

Entry
editThe measles virus has two envelope glycoproteins on the viral surface – hemagglutinin (H) and membrane fusion protein (F). These proteins are responsible for host cell binding and invasion. The H protein mediates receptor attachment and the F protein causes fusion of viral envelope and cellular membrane. Additionally, the F protein can cause infected cells to directly fuse with neighboring uninfected cells forming syncytia. Three receptors for the H protein have been identified to date: complement regulatory molecule CD46, the signaling lymphocyte activation molecule (SLAMF1) and the cell adhesion molecule Nectin-4.[3] For wild type and vaccine strains, extracellular domains of CD150 (SLAM or SLAMF1)[4][5] and/or of nectin-4 (also called Poliovirus-Receptor-Like 4 (PVRL4))[6][7] mainly work as cell entry receptors. A minor fraction of wild type virus strains and all modern vaccine strains derived from the Edmonston strain also use CD46.[8][9]
Genome replication and viral assembly
editOnce the virus has entered a host cell, its strand of negative sense ssRNA is used as a template to create a positive sense copy using the RNA-dependent RNA polymerase that's included in the virion. Then this copy is used to create a new negative copy, and so on, to create many copies of the ssRNA. The positive sense ssRNA is then mass translated by host ribosomes, producing all viral proteins. The viruses are then assembled from their proteins and negative sense ssRNA, and the cell will lyse, discharging the new viral particles and restarting the cycle.[10]

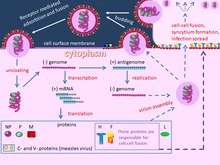
Genome and virion structure
editThe RNA genome of the virus codes 6 main proteins Nucleoprotein (N), Phosphoprotein (P), Matrix protein (M), Fusion protein (F), Hemagglutinin (H), and Large Protein (L),[11] which represents RNA dependent RNA polymerase (RdRp). The viral genome also codes two non-structural proteins C and V. These non-structural proteins are innate immunity antagonists; they help the virus to escape host immune response. Inside the virion genomic RNA is forming complex with N, L and P proteins. N, P and M proteins regulate RNA synthesis by RdRp. The virus is enveloped by a lipid membrane and glycoproteins H and F are virion surface proteins that are associated with this lipid membrane.[citation needed]

Evolution
editThe measles virus evolved from the now eradicated rinderpest virus which infected cattle.[12] Sequence analysis has suggested that the two viruses most probably diverged in the 11th and 12th centuries, though the periods as early as the 5th century fall within the 95% confidence interval of these calculations.[12]
Other analysis has suggested that the divergence may be even older because of the technique's tendency to underestimate ages when strong purifying selection is in action.[13] There is some linguistic evidence for an earlier origin within the seventh century.[11][14] The current epidemic strain evolved at the beginning of the 20th century—most probably between 1908 and 1943.[15]
Genotypes
editThe measles virus genome is typically 15,894 nucleotides long and encodes eight proteins.[16] The WHO currently recognises 8 clades of measles (A–H). Subtypes are designed with numerals—A1, D2 etc. Currently, 23 subtypes are recognised. The 450 nucleotides that code for the C‐terminal 150 amino acids of N are the minimum amount of sequence data required for genotyping a measles virus isolate. The genotyping scheme was introduced in 1998 and extended in 2002 and 2003.[citation needed]
Despite the variety of measles genotypes, there is only one measles serotype. Antibodies to measles bind to the hemagglutinin protein. Thus, antibodies against one genotype (such as the vaccine strain) protect against all other genotypes.[17]
The major genotypes differ between countries and the status of measles circulation within that country or region. Endemic transmission of measles virus was interrupted in the United States and Australia by 2000 and the Americas by 2002.[18]
Infection
editIn the early stages of infection, the measles virus via CD150 (SLAMF1) receptor infects immune cells located in the host respiratory tract such as macrophages and dendritic cells.[19][20][21] They transmit the virus to the lymphoid organs, from which it spreads systemically. In the later stages of infection, the virus infects other immune cell types, including B cells[22] and T lymphocytes[23] also via SLAMF1 receptor. In addition, it infects epithelial cells located in the airways. These cells become infected via nectin-4 receptor and by cell to cell contacts with infected immune cells. The infection of epithelial cells allows the virus to be released via the airstream.[24][25]
References
edit- ^ "ICTV Taxonomy history: Measles morbillivirus". International Committee on Taxonomy of Viruses (ICTV). Retrieved 15 January 2019.
- ^ CDC (5 November 2020). "Measles is Easily Transmitted". Centers for Disease Control and Prevention. Retrieved 28 December 2023.
- ^ Lu G, Gao GF, Yan J (2013). "The receptors and entry of measles virus: a review". Sheng Wu Gong Cheng Xue Bao (in Chinese). 29 (1): 1–9. PMID 23631113.
- ^ Tatsuo, Hironobu; Ono, Nobuyuki; Tanaka, Kotaro; Yanagi, Yusuke (24 August 2000). "SLAM (CDw150) is a cellular receptor for measles virus". Nature. 406 (6798): 893–897. Bibcode:2000Natur.406..893T. doi:10.1038/35022579. ISSN 0028-0836. PMID 10972291. S2CID 4409405.
- ^ Tatsuo, Hironobu; Yanagi, Yusuke (March 2002). "The Morbillivirus Receptor SLAM (CD150)". Microbiology and Immunology. 46 (3): 135–142. doi:10.1111/j.1348-0421.2002.tb02678.x. ISSN 0385-5600. PMID 12008921. S2CID 30651799.
- ^ Mühlebach, Michael D Mateo, Mathieu Sinn, Patrick L Prüfer, Steffen Uhlig, Katharina M Leonard, Vincent H J Navaratnarajah, Chanakha K Frenzke, Marie Wong, Xiao X Sawatsky, Bevan Ramachandran, Shyam Mccray Jr, Paul B Cichutek, Klaus Von Messling, Veronika Lopez, Marc Cattaneo, Roberto. Adherens junction protein nectin-4 is the epithelial receptor for measles virus. OCLC 806252697.
{{cite book}}: CS1 maint: multiple names: authors list (link) - ^ Noyce, Ryan S.; Richardson, Christopher D. (20 June 2012). "Nectin 4 is the epithelial cell receptor for measles virus". Trends in Microbiology. 20 (9): 429–439. doi:10.1016/j.tim.2012.05.006. ISSN 0966-842X. PMID 22721863.
- ^ Erlenhöfer, Christian; Duprex, W. Paul; Rima, Bert K.; ter Meulen, Volker; Schneider-Schaulies, Jürgen (1 June 2002). "Analysis of receptor (CD46, CD150) usage by measles virus". Journal of General Virology. 83 (6): 1431–1436. doi:10.1099/0022-1317-83-6-1431. ISSN 0022-1317. PMID 12029158.
- ^ Lin, Liang-Tzung; Richardson, Christopher (20 September 2016). "The Host Cell Receptors for Measles Virus and Their Interaction with the Viral Hemagglutinin (H) Protein". Viruses. 8 (9): 250. doi:10.3390/v8090250. ISSN 1999-4915. PMC 5035964. PMID 27657109.
- ^ Jiang, Yanliang; Qin, Yali; Chen, Mingzhou (16 November 2016). "Host–Pathogen Interactions in Measles Virus Replication and Anti-Viral Immunity". Viruses. 8 (11): 308. doi:10.3390/v8110308. ISSN 1999-4915. PMC 5127022. PMID 27854326.
- ^ a b Griffin DE (2007). "Measles Virus". In Martin, Malcolm A.; Knipe, David M.; Fields, Bernard N.; Howley, Peter M.; Griffin, Diane; Lamb, Robert (eds.). Fields' virology (5th ed.). Philadelphia: Wolters Kluwer Health/Lippincott Williams & Wilkins. ISBN 978-0-7817-6060-7.
- ^ a b Furuse Y, Suzuki A, Oshitani H (2010). "Origin of measles virus: divergence from rinderpest virus between the 11th and 12th centuries". Virol. J. 7: 52. doi:10.1186/1743-422X-7-52. PMC 2838858. PMID 20202190.
- ^ Wertheim, J. O.; Kosakovsky Pond, S. L. (2011). "Purifying Selection Can Obscure the Ancient Age of Viral Lineages". Molecular Biology and Evolution. 28 (12): 3355–65. doi:10.1093/molbev/msr170. PMC 3247791. PMID 21705379.
- ^ McNeil, W. (1976). Plagues and Peoples. New York: Anchor Press/Doubleday. ISBN 978-0-385-11256-7.
- ^ Pomeroy LW, Bjørnstad ON, Holmes EC (February 2008). "The evolutionary and epidemiological dynamics of the paramyxoviridae". J. Mol. Evol. 66 (2): 98–106. Bibcode:2008JMolE..66...98P. doi:10.1007/s00239-007-9040-x. PMC 3334863. PMID 18217182.
- ^ Phan, MVT; Schapendonk, CME; Oude Munnink, BB; Koopmans, MPG; de Swart, RL; Cotten, M (29 March 2018). "Complete Genome Sequences of Six Measles Virus Strains". Genome Announcements. 6 (13). doi:10.1128/genomeA.00184-18. PMC 5876482. PMID 29599155.
- ^ "Measles". Biologicals — Vaccine Standardization. World Health Organization. 11 March 2013. Archived from the original on 1 May 2014.
- ^ "No endemic transmission of measles in the Americas since 2002". Pan American Health Organization. 14 February 2017. Retrieved 11 September 2017.
- ^ Ferreira, Claudia S. Antunes; Frenzke, Marie; Leonard, Vincent H. J.; Welstead, G. Grant; Richardson, Christopher D.; Cattaneo, Roberto (15 March 2010). "Measles Virus Infection of Alveolar Macrophages and Dendritic Cells Precedes Spread to Lymphatic Organs in Transgenic Mice Expressing Human Signaling Lymphocytic Activation Molecule (SLAM, CD150)". Journal of Virology. 84 (6): 3033–3042. doi:10.1128/JVI.01559-09. ISSN 0022-538X. PMC 2826031. PMID 20042501.
- ^ Leonard, Vincent H.J.; Sinn, Patrick L.; Hodge, Gregory; Miest, Tanner; Devaux, Patricia; Oezguen, Numan; Braun, Werner; McCray, Paul B.; McChesney, Michael B.; Cattaneo, Roberto (1 July 2008). "Measles virus blind to its epithelial cell receptor remains virulent in rhesus monkeys but cannot cross the airway epithelium and is not shed". The Journal of Clinical Investigation. 118 (7): 2448–2458. doi:10.1172/JCI35454. ISSN 0021-9738. PMC 2430500. PMID 18568079.
- ^ Allen, Ingrid V.; McQuaid, Stephen; Penalva, Rosana; Ludlow, Martin; Duprex, W. Paul; Rima, Bertus K. (9 May 2018). "Macrophages and Dendritic Cells Are the Predominant Cells Infected in Measles in Humans". mSphere. 3 (3). doi:10.1128/mSphere.00570-17. ISSN 2379-5042. PMC 5956143. PMID 29743202.
- ^ Laksono, Brigitta M.; Grosserichter-Wagener, Christina; Vries, Rory D. de; Langeveld, Simone A. G.; Brem, Maarten D.; Dongen, Jacques J. M. van; Katsikis, Peter D.; Koopmans, Marion P. G.; Zelm, Menno C. van; Swart, Rik L. de (15 April 2018). "In Vitro Measles Virus Infection of Human Lymphocyte Subsets Demonstrates High Susceptibility and Permissiveness of both Naive and Memory B Cells". Journal of Virology. 92 (8): e00131-18. doi:10.1128/JVI.00131-18. ISSN 0022-538X. PMC 5874404. PMID 29437964.
- ^ Koethe, Susanne; Avota, Elita; Schneider-Schaulies, Sibylle (15 September 2012). "Measles Virus Transmission from Dendritic Cells to T Cells: Formation of Synapse-Like Interfaces Concentrating Viral and Cellular Components". Journal of Virology. 86 (18): 9773–9781. doi:10.1128/JVI.00458-12. ISSN 0022-538X. PMC 3446594. PMID 22761368.
- ^ Ludlow, Martin; Lemon, Ken; de Vries, Rory D.; McQuaid, Stephen; Millar, Emma L.; van Amerongen, Geert; Yüksel, Selma; Verburgh, R. Joyce; Osterhaus, Albert D. M. E.; de Swart, Rik L.; Duprex, W. Paul (April 2013). "Measles Virus Infection of Epithelial Cells in the Macaque Upper Respiratory Tract Is Mediated by Subepithelial Immune Cells". Journal of Virology. 87 (7): 4033–4042. doi:10.1128/JVI.03258-12. ISSN 0022-538X. PMC 3624209. PMID 23365435.
- ^ Singh, Brajesh K.; Li, Ni; Mark, Anna C.; Mateo, Mathieu; Cattaneo, Roberto; Sinn, Patrick L. (1 August 2016). "Cell-to-Cell Contact and Nectin-4 Govern Spread of Measles Virus from Primary Human Myeloid Cells to Primary Human Airway Epithelial Cells". Journal of Virology. 90 (15): 6808–6817. doi:10.1128/JVI.00266-16. ISSN 0022-538X. PMC 4944272. PMID 27194761.
External links
edit- "Measles virus". International Committee on Taxonomy of Viruses.