Hybodontiformes, commonly called hybodonts, are an extinct group of shark-like cartilaginous fish (chondrichthyans) which existed from the late Devonian to the Late Cretaceous. Hybodonts share a close common ancestry with modern sharks and rays (Neoselachii) as part of the clade Euselachii. They are distinguished from other chondrichthyans by their distinctive fin spines and cephalic spines present on the heads of males. An ecologically diverse group, they were abundant in marine and freshwater environments during the late Paleozoic and early Mesozoic, but were rare in open marine environments by the end of the Jurassic, having been largely replaced by modern sharks, though they were still common in freshwater and marginal marine habitats. They survived until the end of the Cretaceous, before going extinct.
| Hybodontiformes Temporal range: Late Devonian to Late Cretaceous
| |
|---|---|

| |
| Fossil and life restoration of Asteracanthus, from the Late Jurassic of Europe | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Chondrichthyes |
| Subclass: | Elasmobranchii |
| Infraclass: | Euselachii |
| Order: | †Hybodontiformes Patterson, 1966 |
| Families | |
|
See text | |
Etymology
editThe term hybodont comes from the Greek word ὕβος or ὑβός meaning hump or hump-backed and ὀδούς, ὀδοντ meaning tooth. This name was given based on their conical compressed teeth.
Taxonomic history
editHybodonts were first described in the nineteenth century based on isolated fossil teeth (Agassiz, 1837). Hybodonts were first separated from living sharks by Zittel (1911).[1] Although historically argued to have a close relationship with the modern shark order Heterodontiformes, this has been refuted.[2] Hybodontiformes are total group-elasmobranchs and the sister group to Neoselachii, which includes modern sharks and rays. Hybodontiformes and Neoselachii are grouped together in the clade Euselachii, to the exclusion of other total-group elasmobranchs like Xenacanthiformes.[3] Hybodonts are divided into a number of families, but the higher level taxonomy of hybodonts, especially Mesozoic taxa, is poorly resolved.[4]
Description
edit.jpg)

The largest hybodonts reached lengths of 2–3 metres (6.6–9.8 ft),[4] while some other hybodonts were much smaller, with adult body lengths of around 25 centimetres (0.82 ft).[5] Hybodonts had a generally robust bodyform. Due to their cartilaginous skeletons usually disintegrating upon death like other chondrichthyans, hybodonts are generally described and identified based on teeth and fin spine fossils, which are more likely to be preserved.[4] Rare partial or complete skeletons are known from areas of exceptional preservation.[6][4][7]

Hybodonts are recognized as having teeth with a prominent cusp which is higher than lateral cusplets.[8] Hybodont teeth are often preserved as incomplete fossils because the base of the tooth is not well attached to the crown.[8] Hybodonts were initially divided into two groups based on their tooth shape.[9] One group had teeth with acuminate cusps that lacked a pulp cavity; these are called osteodont teeth. The other group had a different cusp arrangement and had a pulp cavity, these are called orthodont teeth.[10] For example, the hybodont species Heterophychodus steinmanni have osteodont teeth with vascular canals of dentine which are arranged vertically parallel to each other, also called ‘tubular dentine’.[11] The crowns of these osteodont teeth are covered with a single layer of enameloid. Hybodont teeth served a variety of functions depending on the species, including grinding, crushing (durophagy), tearing, clutching, and even cutting.[4]
Hybodonts are characterized by having two dorsal fins each preceded by a fin spine. The fin spine morphology is unique to each hybodont species. The fin spines are elongate and gently curved towards the rear, with the posterior part of the spine being covered in hooked denticles, typically in two parallel rows running along the length of the spine, sometimes with a ridge between them. Part of the front of the spines are often covered in a ribbed ornamentation, while in some other hybodonts this region is covered in rows of small bumps. The spines are mineralised, and primary composed of osteodentine, while the ornamentation is formed of enamel.[12] Similar fin spines are also found in many extinct chondrichthyan groups as well as in some modern sharks like Heterodontus and squalids.[13] Male hybodonts had either one or two pairs of cephalic spines on their heads, a characteristic distinctive to hybodonts.[14] These spines, while of variable placement,[10] were always placed posterior to the eye socket,[15] and were composed of a base divided into three lobes, with the main part of the spine being backwardly curved, most specimens of which had a barb near the apex.[10] These spines, like the fin spines, were mineralised, with the base composed of osteodentine,[16] while the main part of the spine was covered in enamel. Male hybodonts possessed fin claspers used in mating, like modern sharks.[10] Hybodonts had a fully heterocercal tail fin, where the upper lobe of the fin was much larger than the lower one due to the spine extending into it.[17] Like living sharks and rays, the skin of hybodonts was covered with dermal denticles.[18] Hybodonts laid egg cases, similar to those produced by living cartilaginous fish. Most hybodont egg cases are assigned to the genus Palaeoxyris, which tapers towards both ends, with one end having a tendril which attached to substrate, with the middle section being composed of at least three twisted bands.[19]
-
 Teeth of Durnovariaodus, a member of the family Hybodontidae
Teeth of Durnovariaodus, a member of the family Hybodontidae -
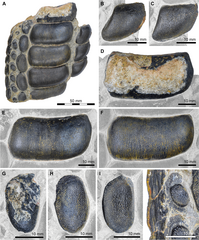 Jaw fragment of Strophodus a specialised durophagous hybodont
Jaw fragment of Strophodus a specialised durophagous hybodont -
 Jaws of Asteracanthus, showing the arrangement of the teeth in jaws. The teeth were designed for grasping
Jaws of Asteracanthus, showing the arrangement of the teeth in jaws. The teeth were designed for grasping -

-
 Teeth of Planohybodus, a hybodontid, whose teeth were designed for tearing
Teeth of Planohybodus, a hybodontid, whose teeth were designed for tearing -
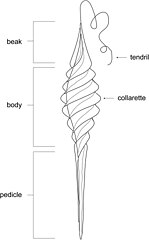 Diagram of Palaeoxyris egg cases, thought to have been produced by hybodonts
Diagram of Palaeoxyris egg cases, thought to have been produced by hybodonts



.jpg)


Ecology
edit
Hybodont fossils are found in depositional environments ranging from marine to fluvial (river deposits).[20] Many hybodonts are thought to have been euryhaline, able to tolerate a wide range of salinities.[21] Hybodonts inhabited freshwater environments from early in their evolutionary history, spanning from the Carboniferous onwards.[22] Based on isotopic analysis, some species of hybodonts are likely to have permanently lived in freshwater environments,[23][24] while others may have migrated between marine and freshwater environments.[25] One genus of hybodont, Onychoselache of the lower Carboniferous of Scotland, is suggested to have been capable of amphibious locomotion, similar to modern orectolobiform sharks such as bamboo and epaulette sharks, due to its well-developed pectoral fins.[16] It has been suggested that male hybodonts used their cephalic spines to grip females during mating.[26] Preserved egg cases of hybodonts assigned to Palaeoxyris indicate that at least some hybodonts laid their eggs in freshwater and brackish environments, with the eggs being attached to vegetation via a tendril. Laying of eggs in freshwater is not known in any living cartilaginous fish.[27][19][28] At least some hybodonts are suggested to have utlilized specific sites as nurseries, such as in the Triassic lake deposits of the Madygen Formation of Kyrgyzstan, where eggs of Lonchidion are suggested to have been laid on the lakeshore or upriver areas, where the juveniles hatched and matured, before migrating deeper into the lake as adults.[27]

Hybodonts are thought to have been generally relatively slow swimmers, though capable of fast bursts of locomotion.[29] Some hybodonts like Hybodus are thought to have been active predators capable of feeding on swiftly moving prey,[2] with preserved stomach contents of a specimen of Hybodus hauffianus indicating that they fed on belemnites (a type of extinct squid-like cephalopod).[30] Hybodonts have a wide variety of tooth shapes. This variety suggests that they took advantage of multiple food sources.[8] It is thought that some hybodonts which had wider, flatter, teeth specialized in crushing or grinding hard-shelled prey (durophagy),[20] with some hybodonts like Asteracanthus probably consuming both hard and soft bodied prey.[4] Often multiple species of hybodonts with different prey preferences coexisted within the same ecosystem.[31][11]
Evolutionary history
edit
The earliest hybodont remains are from the latest Devonian (Famennian, ~ 360 million years ago) of Iran, belonging to the genus Roongodus,[32] as well as remains assigned to Lissodus of the same age from Belgium.[33] Carboniferous hybodonts include both durophagous and non-durophagous forms, while durophagous forms were dominant during the Permian period.[33] By the Permian period, hybodonts had a global distribution.[33][34][35] The Permian-Triassic extinction event only had a limited effect on hybodont diversity.[29] Maximum hybodont diversity is observed during the Triassic. During the Triassic and Early Jurassic, hybodontiforms were the dominant elasmobranchs in both marine and non-marine environments.[20] A shift in hybodonts was seen during the Middle Jurassic, a transition between the distinctly different assemblages seen in the Triassic – Early Jurassic and the Late Jurassic – Cretaceous.[20] As neoselachians (group of modern sharks) diversified further during the Late Jurassic, hybodontiforms became less prevalent in open marine conditions but remained diverse in fluvial and restricted settings during the Cretaceous.[20] Possible reasons for the replacement of hybodonts by modern sharks include more effective locomotory and jaw movement mechanisms of the latter group.[36] By the end of the Cretaceous, hybodonts had declined to only a handful of species,[37] including members of Lonchidion[38], and Meristodonoides.[39] The last hybodonts disappeared, seemingly abruptly, as part of the Cretaceous-Paleogene extinction event approximately 66 million years ago.[37]
Families and genera
editThe taxonomy of hybodonts is considered poorly resolved,[4] so the classification presented should not be taken as authoritative.
- Lonchidiidae Herman, 1977[40]
- Distobatidae
- Acrodontidae
- Hybodontidae
- Incertae sedis
- Tribodus
- Strophodus
- Asteracanthus
- Roongodus
- Polyacrodus
- Palaeobates
- Bdellodus
- Thaiodus
- Acrorhizodus
- Khoratodus
- Arctacanthus
- Reesodus
- Steinbachodus
- Onychoselache
- Omanoselache
- Pororhiza
- Mukdahanodus
- Secarodus
- Hamiltonichthys
- Gansuselache
- Dabasacanthus
- Teresodus
- Diablodontus
- Gunnellodus?
- Heteroptychodus
- Lissodus
- Columnaodus
- Carinacanthus
- Form genera
- Palaeoxyris (genus used for the egg capsules of hybodonts)
References
edit- ^ Zittel, K. von, 1911, Grunzuege der Palaontologie, 2 ed. II. Abt. Vertebrata, vii + 598 pp. R. Oldenburg Verlag, Muchen, Berlin.
- ^ a b Maisey, J. G., 2012, What is an ‘elasmobranch’? The impact of palaeontology in understanding elasmobranch phylogeny and evolution: Journal of Fish Biology, v. 80, no. 5, p. 918-951.
- ^ Coates, Michael I.; Tietjen, Kristen (March 2017). "The neurocranium of the Lower Carboniferous shark Tristychius arcuatus (Agassiz, )". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 108 (1): 19–35. Bibcode:2017EESTR.108...19C. doi:10.1017/S1755691018000130. ISSN 1755-6910. S2CID 135297534.
- ^ a b c d e f g Stumpf, Sebastian; López-Romero, Faviel A.; Kindlimann, René; Lacombat, Frederic; Pohl, Burkhard; Kriwet, Jürgen (August 2021). Cavin, Lionel (ed.). "A unique hybodontiform skeleton provides novel insights into Mesozoic chondrichthyan life". Papers in Palaeontology. 7 (3): 1479–1505. Bibcode:2021PPal....7.1479S. doi:10.1002/spp2.1350. ISSN 2056-2799. S2CID 234204226.
- ^ Vullo, Romain; Néraudeau, Didier; Dépré, Eric (October 2013). "Vertebrate remains from the Cenomanian (Late Cretaceous) plant-bearing Lagerstätte of Puy-Puy (Charente-Maritime, France)". Cretaceous Research. 45: 314–320. Bibcode:2013CrRes..45..314V. doi:10.1016/j.cretres.2013.06.002.
- ^ Lane, J. A., and Maisey, J. G., 2009, Pectoral Anatomy of Tribodus limae (Elasmobranchii: Hybodontiformes) from the Lower Cretaceous of Northeastern Brazil: Journal of Vertebrate Paleontology, v. 29, no. 1, p. 25-38.
- ^ Wang, N.-Z.; Zhang, X.; Zhu, M.; Zhao, W.-J. (May 2009). "A new articulated hybodontoid from Late Permian of northwestern China". Acta Zoologica. 90: 159–170. doi:10.1111/j.1463-6395.2008.00382.x.
- ^ a b c Koot, M. B., Cuny, G., Tintori, A., and Twitchett, R. J., 2013, A new diverse shark fauna from the Wordian (Middle Permian) Khuff Formation in the interior Haushi-Huqf area, Sultanate of Oman: Palaeontology, v. 56, no. 2, p. 303-343.
- ^ Agassiz, L., 1833-1844, Recherches sur les poisons fossils. Neuchatel, 5 vols. 1420 pp. with supplement.
- ^ a b c d Maisey, J. G., 1982, The anatomy and interrelationships of Mesozoic hybodont sharks: American Museum Novitates, v. 2724.
- ^ a b Cuny, G., Suteethorn, V., Buffetaut, E., and Philippe, M., 2003, Hybodont sharks from the Mesozoic Khorat Group of Thailand: Mahasarakham University Journal, v. 22.
- ^ Maisey, J. G., 1978, Growth and form of spines in hybodont sharks: Palaeontology, v. 21, no. 3, p. 657-666.
- ^ Maisey, John G. (2009-03-12). "The spine-brush complex in symmoriiform sharks (Chondrichthyes; Symmoriiformes), with comments on dorsal fin modularity". Journal of Vertebrate Paleontology. 29 (1): 14–24. Bibcode:2009JVPal..29...14M. doi:10.1671/039.029.0130. ISSN 0272-4634. S2CID 86408226.
- ^ Rees, Jan (March 2002). "Shark fauna and depositional environment of the earliest Cretaceous Vitabäck Clays at Eriksdal, southern Sweden". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 93 (1): 59–71. doi:10.1017/S0263593300000328. ISSN 1473-7116. S2CID 130984933.
- ^ Stumpf, Sebastian; Etches, Steve; Underwood, Charlie J.; Kriwet, Jürgen (2021-05-11). "Durnonovariaodus maiseyi gen. et sp. nov., a new hybodontiform shark-like chondrichthyan from the Upper Jurassic Kimmeridge Clay Formation of England". PeerJ. 9: e11362. doi:10.7717/peerj.11362. ISSN 2167-8359. PMC 8121075. PMID 34026354.
- ^ a b Coates, Michael I.; Gess, Robert W. (2007). "A New Reconstruction of Onychoselache Traquairi, Comments on Early Chondrichthyan Pectoral Girdles and Hybodontiform Phylogeny". Palaeontology. 50 (6): 1421–1446. Bibcode:2007Palgy..50.1421C. doi:10.1111/j.1475-4983.2007.00719.x. S2CID 140556654.
- ^ Kim, Sun H.; Shimada, Kenshu; Rigsby, Cynthia K. (March 2013). "Anatomy and Evolution of Heterocercal Tail in Lamniform Sharks: CAUDAL FINS OF LAMNIFORM SHARKS". The Anatomical Record. 296 (3): 433–442. doi:10.1002/ar.22647. PMID 23381874. S2CID 205411092.
- ^ Maisey, John G.; Denton, John S. S. (2016-09-02). "Dermal denticle patterning in the Cretaceous hybodont shark Tribodus limae (Euselachii, Hybodontiformes), and its implications for the evolution of patterning in the chondrichthyan dermal skeleton". Journal of Vertebrate Paleontology. 36 (5): e1179200. Bibcode:2016JVPal..36E9200M. doi:10.1080/02724634.2016.1179200. ISSN 0272-4634. S2CID 88591830.
- ^ a b Fischer, Jan; Axsmith, Brian J.; Ash, Sidney R. (2010-03-01). "First unequivocal record of the hybodont shark egg capsule Palaeoxyris in the Mesozoic of North America". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 255 (3): 327–344. doi:10.1127/0077-7749/2009/0028. ISSN 0077-7749.
- ^ a b c d e Rees, J. A. N., and Underwood, C. J., 2008, Hybodont sharks of the English Bathonian and Callovian (Middle Jurassic): Palaeontology, v. 51, no. 1, p. 117-147.
- ^ Batchelor, Trevor John (December 2013). "A new species of Vectiselachos (Chondrichthyes, Selachii) from the Early Cretaceous of southern England". Proceedings of the Geologists' Association. 124 (6): 967–972. Bibcode:2013PrGA..124..967B. doi:10.1016/j.pgeola.2013.05.001.
- ^ Wen, Wen; Kriwet, JüRgen; Zhang, Qiyue; Benton, Michael J.; Duffin, Christopher J.; Huang, Jingyuan; Zhou, Changyong; Hu, Shixue; Ma, Zhixin (2022-06-30). "Hybodontiform shark remains (Chondrichthyes, Elasmobranchii) from the Lower Triassic of Yunnan Province, China, with comments on hybodontiform diversity across the PTB". Journal of Vertebrate Paleontology. 42 (1). Bibcode:2022JVPal..42E8712W. doi:10.1080/02724634.2022.2108712. ISSN 0272-4634. S2CID 252136919.
- ^ Fischer, Jan; Schneider, Jörg W.; Voigt, Silke; Joachimski, Michael M.; Tichomirowa, Marion; Tütken, Thomas; Götze, Jens; Berner, Ulrich (2013-03-29). "Oxygen and strontium isotopes from fossil shark teeth: Environmental and ecological implications for Late Palaeozoic European basins". Chemical Geology. 342: 44–62. Bibcode:2013ChGeo.342...44F. doi:10.1016/j.chemgeo.2013.01.022. ISSN 0009-2541.
- ^ Klug, Stefanie; Tütken, Thomas; Wings, Oliver; Pfretzschner, Hans-Ulrich; Martin, Thomas (2010-09-01). "A Late Jurassic freshwater shark assemblage (Chondrichthyes, Hybodontiformes) from the southern Junggar Basin, Xinjiang, Northwest China". Palaeobiodiversity and Palaeoenvironments. 90 (3): 241–257. Bibcode:2010PdPe...90..241K. doi:10.1007/s12549-010-0032-2. ISSN 1867-1608. S2CID 129236098.
- ^ Leuzinger, L.; Kocsis, L.; Billon-Bruyat, J.-P.; Spezzaferri, S.; Vennemann, T. (2015-12-07). "Stable isotope study of a new chondrichthyan fauna (Kimmeridgian, Porrentruy, Swiss Jura): an unusual freshwater-influenced isotopic composition for the hybodont shark <i>Asteracanthus</i>". Biogeosciences. 12 (23): 6945–6954. Bibcode:2015BGeo...12.6945L. doi:10.5194/bg-12-6945-2015. hdl:11336/80921. ISSN 1726-4189. S2CID 129044300.
- ^ Turmine-Juhel, Pernelle; Wilks, Richard; Brockhurst, David; Austen, Peter A.; Duffin, Christopher J.; Benton, Michael J. (December 2019). "Microvertebrates from the Wadhurst Clay Formation (Lower Cretaceous) of Ashdown Brickworks, East Sussex, UK". Proceedings of the Geologists' Association. 130 (6): 752–769. Bibcode:2019PrGA..130..752T. doi:10.1016/j.pgeola.2019.08.003. S2CID 202904040.
- ^ a b Fischer, J. A. N., Voigt, S., Schneider, J. W., Buchwitz, M., and Voigt, S., 2011, A selachian freshwater fauna from the Triassic of Kyrgyzstan and its implication for Mesozoic shark nurseries: Journal of Vertebrate Paleontology, v. 31, no. 5, p. 937-953.
- ^ Krüger, Ashley; Slater, Sam; Vajda, Vivi (2021-07-03). "3D imaging of shark egg cases ( Palaeoxyris ) from Sweden with new insights into Early Jurassic shark ecology". GFF. 143 (2–3): 229–247. Bibcode:2021GFF...143..229K. doi:10.1080/11035897.2021.1907442. ISSN 1103-5897.
- ^ a b Wen, Wen; Zhang, Qiyue; Benton, Michael J.; Kriwet, Jürgen; Hu, Shixue; Huang, Jinyuan; Zhou, Changyong; Cui, Xindong; Ma, Zhixin; Min, Xiao (February 2023). "First occurrence of hybodontid teeth in the Luoping Biota (Middle Triassic, Anisian), emphasizing recovery of the marine ecosystem after the end-Permian mass extinction". Palaeogeography, Palaeoclimatology, Palaeoecology. 617: 111471. doi:10.1016/j.palaeo.2023.111471.
- ^ Klug, Christian; Schweigert, Günter; Hoffmann, René; Weis, Robert; De Baets, Kenneth (December 2021). "Fossilized leftover falls as sources of palaeoecological data: a 'pabulite' comprising a crustacean, a belemnite and a vertebrate from the Early Jurassic Posidonia Shale". Swiss Journal of Palaeontology. 140 (1): 10. Bibcode:2021SwJP..140...10K. doi:10.1186/s13358-021-00225-z. ISSN 1664-2376. PMC 8549986. PMID 34721282.
- ^ Cappetta, H., Buffetaut, E., Cuny, G., and Suteethorn, V., 2006, A new Elasmobranch assemblage from the Lower Cretaceous of Thailand Palaeontology, v. 49, no. 3, p. 547-555.
- ^ Hairapetian, V. and Ginter, M. 2009. Famennian chondrichthyan remains from the Chahriseh section, central Iran. Acta Geologica Polonica, 59, 173–200.
- ^ a b c Hodnett, J-P., Elliott, D. K., and Olson, T. J. 2013. A new basal hybodont (Chondrichthyes, Hybodontiformes) from the Middle Permian (Roadian) Kaibab Formation, of northern Arizona. New Mexico Museum of Natural History and Science Bulletin, 60:103–108.
- ^ Peecook, Brandon R.; Bronson, Allison W.; Otoo, Benjamin K.A.; Sidor, Christian A. (November 2021). "Freshwater fish faunas from two Permian rift valleys of Zambia, novel additions to the ichthyofauna of southern Pangea". Journal of African Earth Sciences. 183: 104325. Bibcode:2021JAfES.18304325P. doi:10.1016/j.jafrearsci.2021.104325.
- ^ Cione, Alberto Luis; Gouiric-Cavalli, Soledad; Mennucci, Jorge Augusto; Cabrera, Daniel Alfredo; Freije, Rubén Hugo (2010). "First vertebrate body remains from the Permian of Argentina (Elasmobranchii and Actinopterygii)". Proceedings of the Geologists' Association. 121 (3): 301–312. Bibcode:2010PrGA..121..301C. doi:10.1016/j.pgeola.2010.04.003. hdl:11336/242890.
- ^ Sternes, Phillip C.; Schmitz, Lars; Higham, Timothy E. (June 2024). "The rise of pelagic sharks and adaptive evolution of pectoral fin morphology during the Cretaceous". Current Biology. 34 (12): 2764–2772.e3. doi:10.1016/j.cub.2024.05.016. PMID 38834065.
- ^ a b Kriwet, Jürgen; Benton, Michael J. (November 2004). "Neoselachian (Chondrichthyes, Elasmobranchii) diversity across the Cretaceous–Tertiary boundary". Palaeogeography, Palaeoclimatology, Palaeoecology. 214 (3): 181–194. doi:10.1016/j.palaeo.2004.02.049.
- ^ Gates, Terry A.; Gorscak, Eric; Makovicky, Peter J. (May 2019). "New sharks and other chondrichthyans from the latest Maastrichtian (Late Cretaceous) of North America". Journal of Paleontology. 93 (3): 512–530. Bibcode:2019JPal...93..512G. doi:10.1017/jpa.2018.92. ISSN 0022-3360.
- ^ Stumpf, Sebastian; Meng, Stefan; Kriwet, Jürgen (2022-01-26). "Diversity Patterns of Late Jurassic Chondrichthyans: New Insights from a Historically Collected Hybodontiform Tooth Assemblage from Poland". Diversity. 14 (2): 85. doi:10.3390/d14020085. ISSN 1424-2818.
- ^ Manzanares, Esther; Pla, Cristina; Martínez-Pérez, Carlos; Ferrón, Humberto; Botella, Héctor (2017-01-02). "Lonchidion derenzii , sp. nov., a new lonchidiid shark (Chondrichthyes, Hybodontiforms) from the Upper Triassic of Spain, with remarks on lonchidiid enameloid". Journal of Vertebrate Paleontology. 37 (1): e1253585. Bibcode:2017JVPal..37E3585M. doi:10.1080/02724634.2017.1253585. hdl:10550/85565. ISSN 0272-4634. S2CID 132467956.